为什么光合作用的过程不能被机器模仿?
这是我熟悉的领域。
我们农学的《生物化学》课里有对光合作用在分子层面和电子层面的详细解答,我们一步一步地来看看能否把光合作用进行机械化模仿吧。
先照顾一下小学生,讲一下概念。
光合作用是糖类合成代谢的主要途径,糖类又可以进一步转化成蛋白质和脂类,这三类物质构成了地球生命体的物质基础。
光合作用是指绿色植物、光合细菌或藻类等以二氧化碳为碳源,水为供氢体,利用叶绿素分子捕获的光能为能源,合成以糖类物质为主的有机化合物,同时释放出氧气的过程。这一过程的最终结果是将光能变成了化学能。
光合作用的总反应式可表示如下:
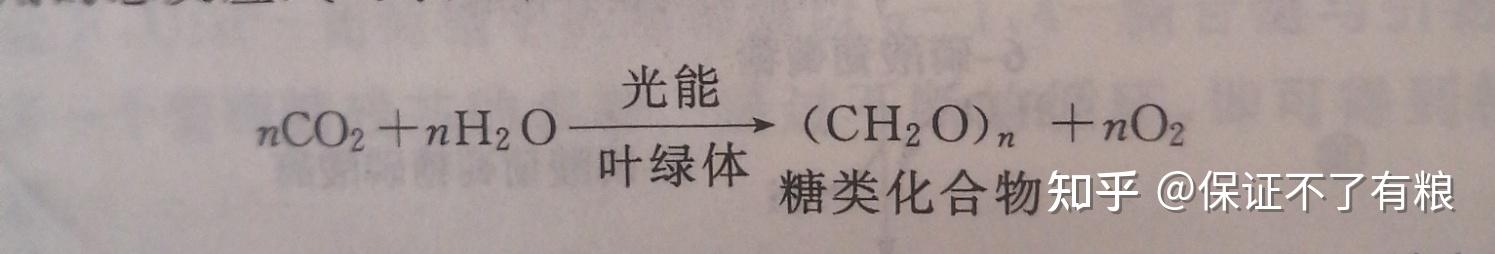
糖类化合物是人、动物和大多数细菌所利用的最重要能源。从根本上讲,太阳能是地球上所有代谢能量的最终来源。光合作用是生物界规模最大的生物化学过程。
叶绿体
植物的绿色部分含有叶绿体,叶绿体内含有叶绿素等光合色素,是绿色植物进行光合作用的场所。
带颜色的长这样:

不带颜色的长这样:
总之,我觉得它很酷很科幻,是地球生命的能量工厂。
如果人类要造一套可以进行光合作用的机器的话,估计也只能造成这个形状。
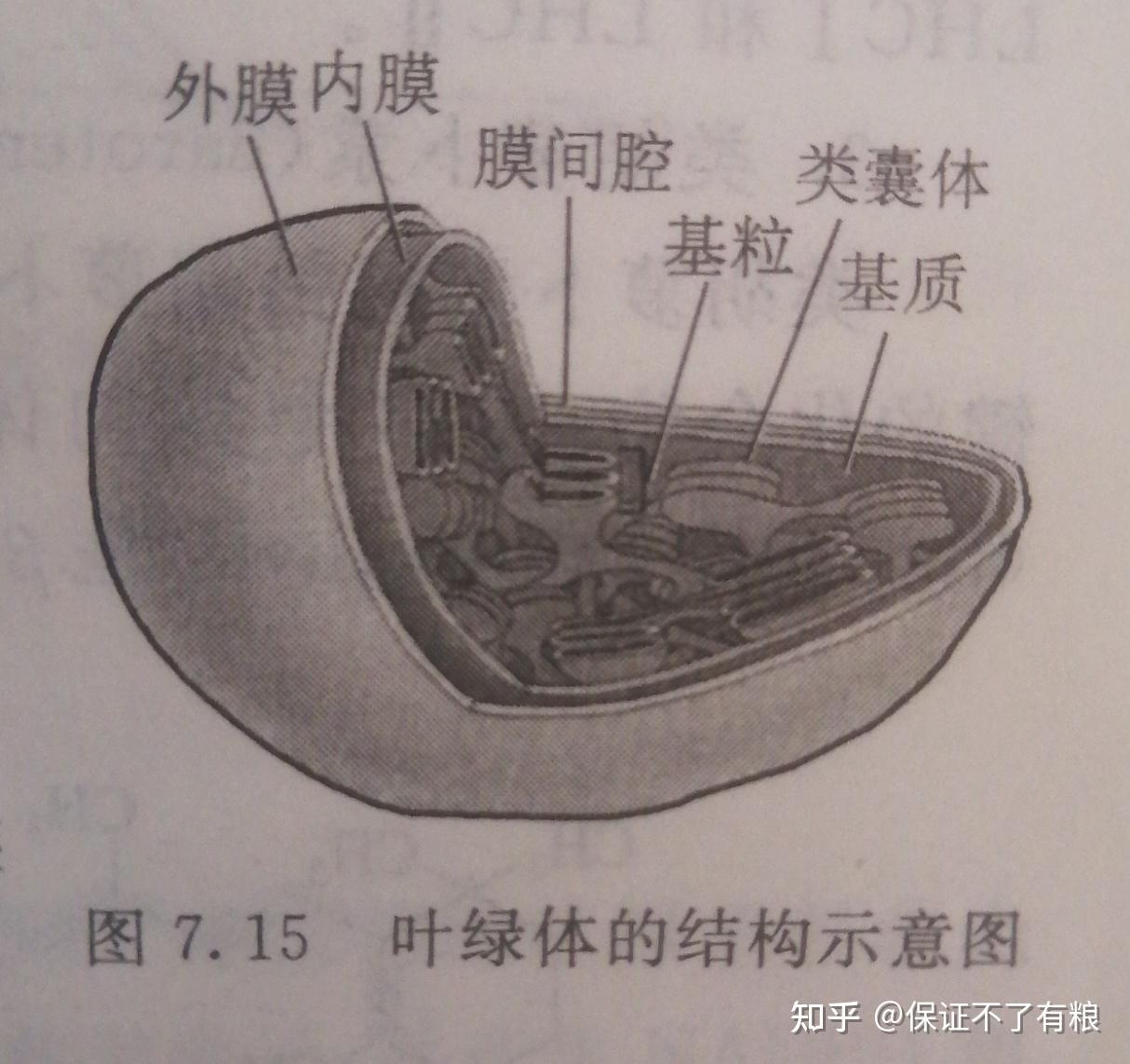
叶绿体由外膜和内膜组成,内外膜之间有间隙。如图7.15,叶绿体内膜内为基质,包含有各种可溶性酶、核糖体以及环状 DNA 分子等,是进行暗反应的场所。基质内还分布着具有膜结构特点的片层状类囊体。片层状类囊体在有些部位堆叠成颗粒状,称为基粒。类囊体含有大量可进行光反应的光合色素。
这里的可溶性酶、核糖体、环状DNA分子,可能是机器进行光合作用的耗材。
光合细菌无叶绿体,它们的光合色素存在于类似的片层结构中。
叶绿素
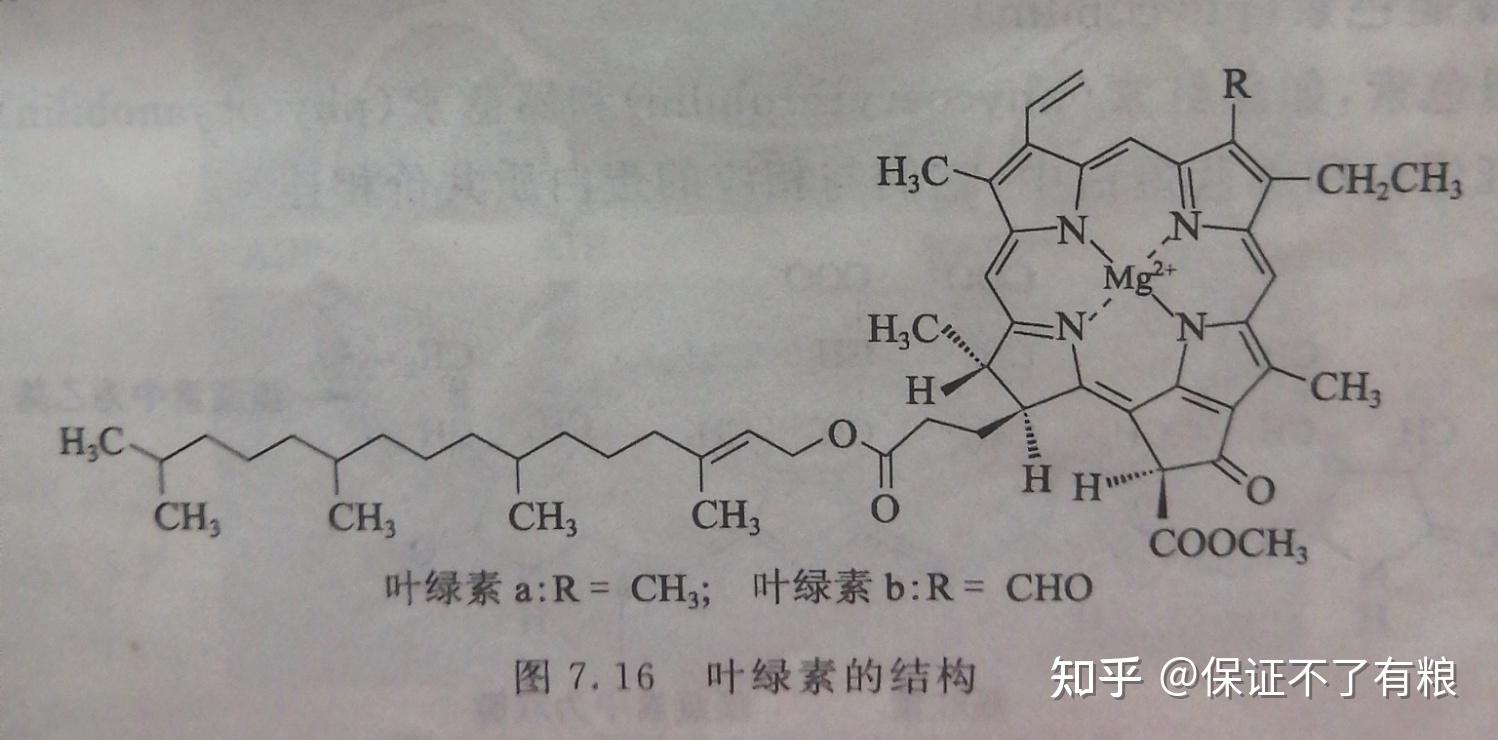
绿色植物叶绿体中接受光能的主要组分是叶绿素,包括叶绿素 a 和叶绿素 b 。其他的光合色素,如类胡萝卜素等,作为辅助色素在光合作用中处于次要地位。光合细菌和藻类中还含有叶绿素 c 和藻胆色素等。
叶绿素是一类含镁的卟咻衍生物,其镁卟咻环带羧基的侧链与一个含有20个碳的植醇形成酯。叶绿素 a 与叶绿素 b 之间的差别在于吡咯环上的一个基团不同(图7.16)。
以我们现在的生物化学技术,这种分子生产出来应该不是很难。
叶绿素不溶于水,能溶于有机溶剂。叶绿素分子具有两亲性(既亲水又亲脂)的特点,亲水的镁卟琳"头"能插入蛋白质层,亲脂的植醇"尾"则深入到内膜的磷脂双分子层中。叶绿素分子是一个大的共轭体系,在可见光区有很强的吸收。
蛋白质层和磷脂双分子层也不难制造出来,不过,由于人造机器的体积注定会是肉眼可见级别的大,用什么来支撑和保护这么精细到分子厚度的膜倒是一个工程学难题,在精微生物工程学上,我们的经验并不足够丰富。
不同的叶绿素分子,它们的特征吸收也不相同:叶绿素 a 和叶绿素 b 的最大吸收波长分别为680nm和460 nm 。固体状态的叶绿素呈黑色,溶液状态的叶绿素则呈绿色,其中叶绿素 a 呈蓝绿色,叶绿素 b 呈黄绿色。
在高等植物和藻类中,叶绿素与蛋白质形成复合体,以缀合蛋白质的形式存在于光合膜(类囊体膜)中,称为集光复合体。它是光系统的天线,因此又将这种叶绿素称为天线色素。每种光系统有其相应的集光复合体,如光系统1和光系统2的集光复合体分别是 LHC1和 LHC2。
天线下面还有一大坨处理器件?
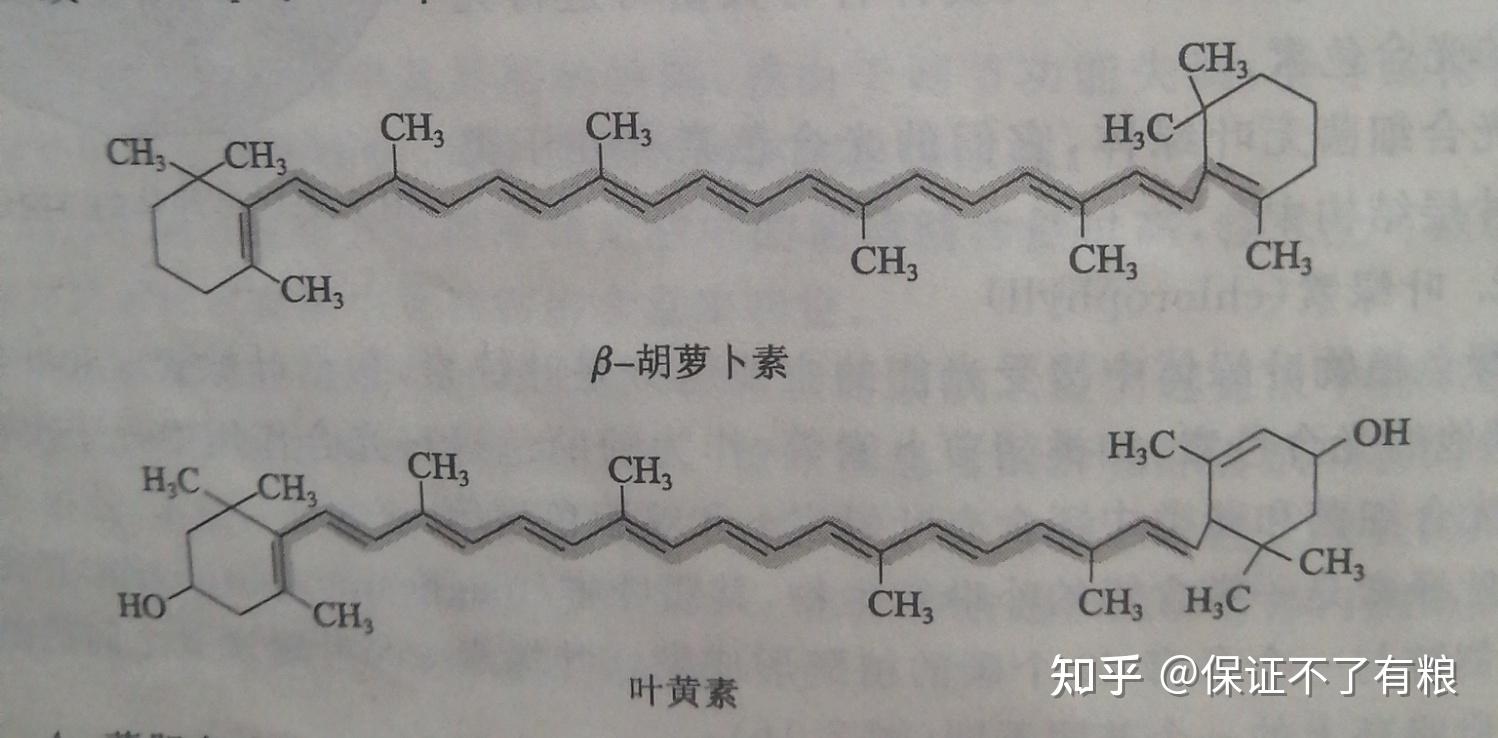
类胡萝卜素~辅助色素
类胡萝卜素包括胡萝卜素和叶黄素等。胡萝卜素是一个含有11个共轭双健的化合物,有多个异构体,常见的是贝塔﹣胡萝卜素。叶黄素是 贝塔﹣胡萝卜素衍生的二元醇。它们的结构如下:
藻胆色素这种赵姨娘角色就不讲了,留点心力看后面。
各种光合色素分子的结构中都存在大的共轭体系,因此它们能够吸收可见光,而且特征吸收波长各不相同,如图7.17,因此可以产生各种各样的颜色。
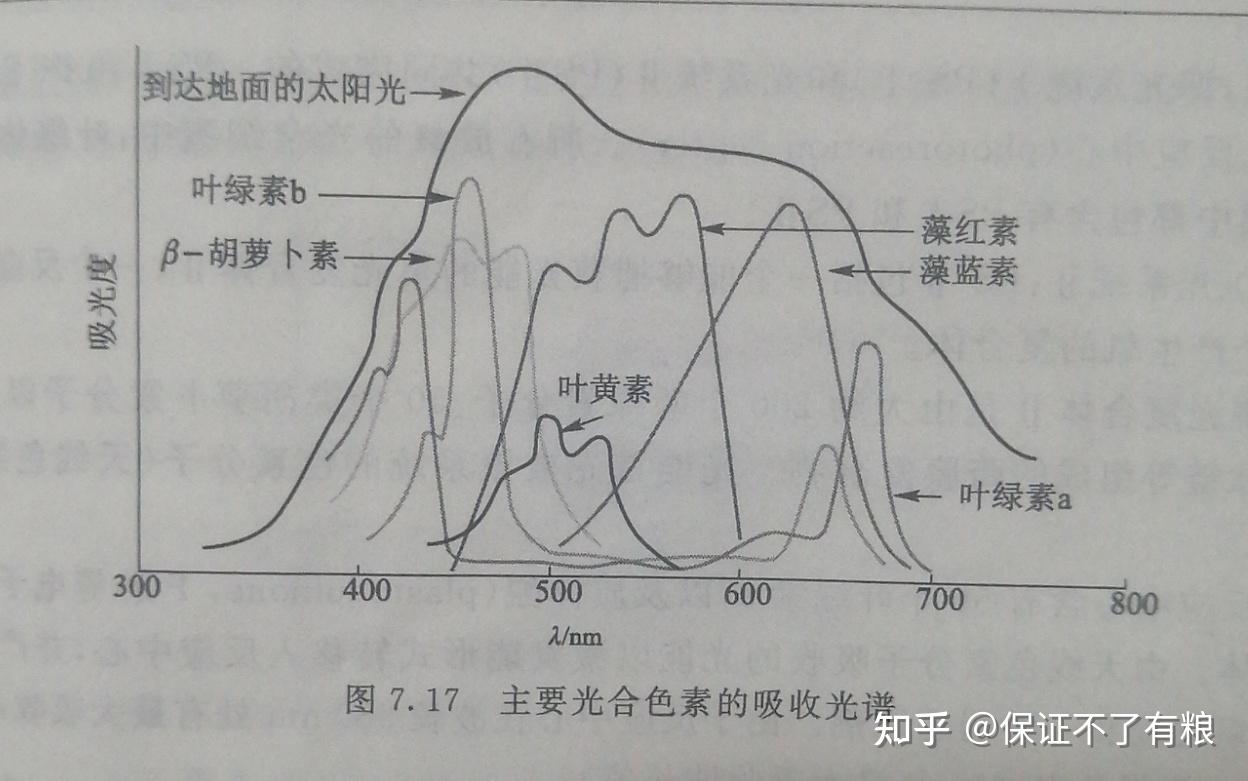
这也是色彩斑斓的自然界形成的主要原因,光合色素分子把红橙黄绿青蓝紫中的一些颜色光吸收了,白光不能再合一成白光,自然物就将那些剩下的颜色光反射给我们的眼睛,于是,我们就看到了这色彩斑斓的世界。
光合作用机制
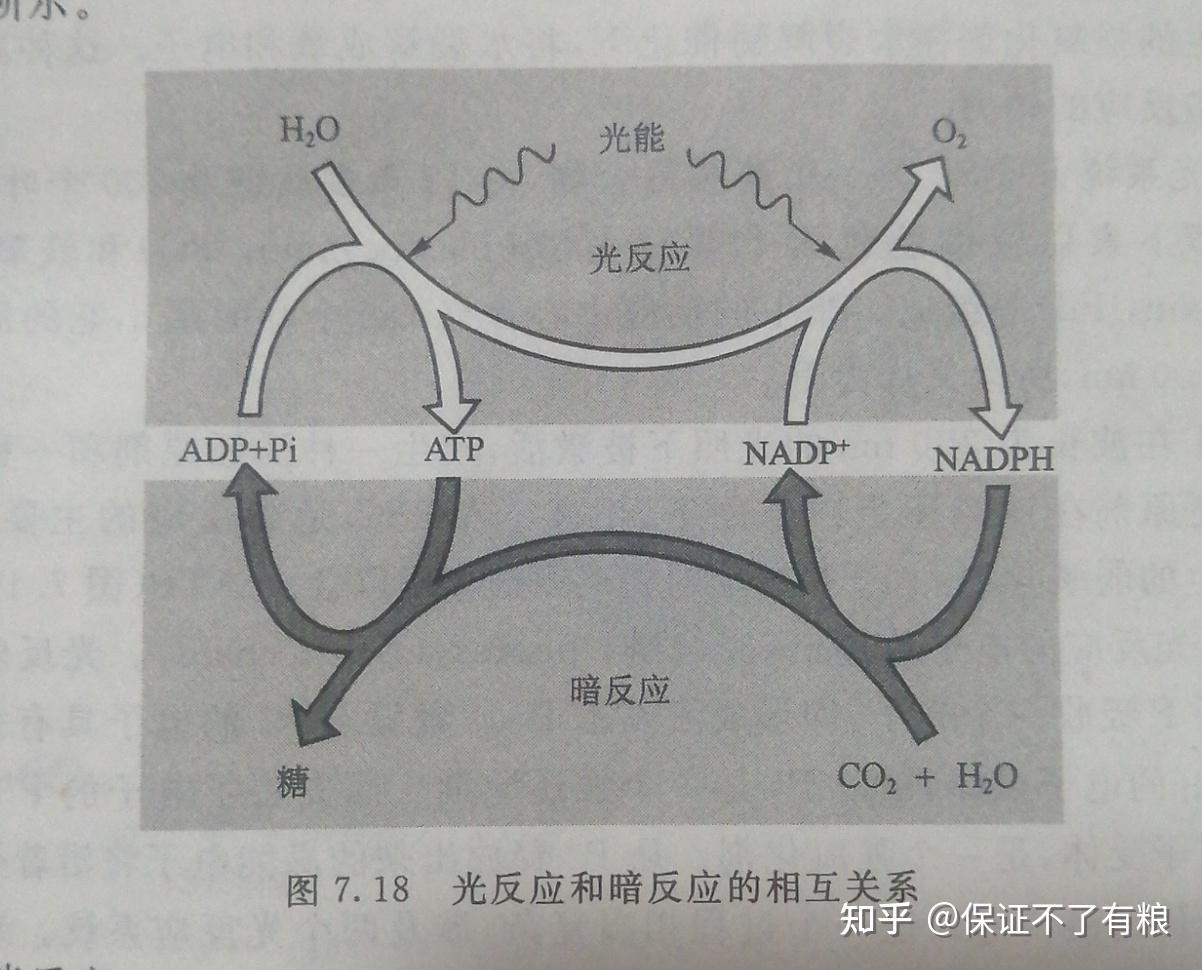
绿色植物的光合作用由光反应和暗反应组成。
光反应( light reaction )是光能转变成化学能的反应,即植物的叶绿素吸收光能进行光化学反应,使水分子活化裂解出O2, H +和释放出电子,并产生 NADPH 和 ATP 。暗反应( dark reaction )为酶促反应,由光反应产生的 NADPH 在 ATP 供给能量的条件下,使CO2还原成简单糖类。这两类反应在光合作用中的相互关系如图7.18所示。
光反应
(1)光反应系统( photosystem , PS )。光反应过程是由含叶绿素的两种光合系统,即光系统 I ( PSI )和光系统 II ( PSII )共同完成的。 PSI 和 PSII 又被称为光反应中心( photoreaction center )。所有放氧的光合细胞中,叶绿体的类囊体膜中都包含有 PSI 和 PSII 。
①光系统 II : PS II 包括一个能够捕获光能的集光复合体 II、一个反应中心及一个产生氧的复合体。
集光复合体 II 是由大约200个叶绿素分子、50个类胡萝卜素分子以及12条多肽链等组成的跨膜复合物。光能首先被该系统的色素分子(天线色素)所吸收。
反应中心含有50个叶绿素 a ,以及质体醌( plastoquinone , PQ )等电子供体和受体。由天线色素分子吸收的光能以激发能形式转移入反应中心,并产生一种强氧化剂和一种弱还原剂。由于反应中心在波长680nm处有最大吸收,又称为P680( P 指色素,680是最大吸收波长值)。
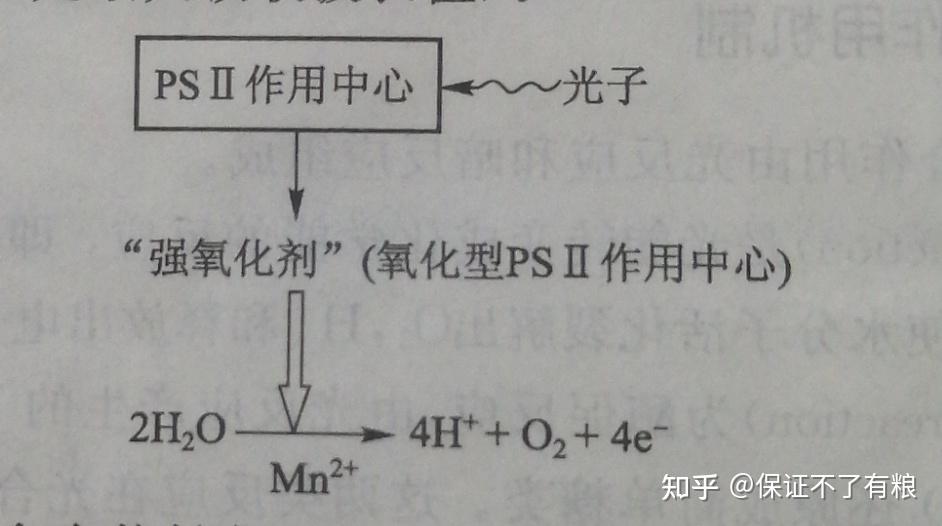
产生氧的复合体含有能够促进水裂解的蛋白质(含有 Mn +离子)等。反应中心产生的强氧化剂在水裂解酶催化下,将水裂解成氧和电子。这种高能电子是推动暗反应的动力。
这个系统不容易做噢,而且这句“大约200个叶绿素分子、50个类胡萝卜素分子以及12条多肽链等”里有个等,这一句“以及质体醌( plastoquinone , PQ )等电子供体和受体”里也有一个等,“等”里面的物质肯定不是最多的,但很有可能是秘方成分,没有这样的成分和成分之间严格的数量配比,光能就不一定能变成激发能形式,也就不一定能转移入反应中心,后面过程中要用到的强氧化剂和弱还原剂也不一定能够产生。
让我们接着往下看生物的复杂性。
②光系统 I : PSI 是一个跨膜复合物,由13条多肽链及200个叶绿素、50个类胡萝卜素以及细胞色素 f 、质体蓝素( plastocyanin , PC )和铁氧还蛋白( ferredoxin , Fd )等组成。 PSI 的反应中心含有130个叶绿素 a ,它的最大吸收波长为700nm,所以又称为P700。
PSI 在波长为700nm的光照下被激活,产生一种强还原剂和一种弱氧化剂。强还原剂在铁氧还蛋白作用下,生成 NADPH ,是暗反应的主要还原剂。 PSI 产生的弱氧化剂和 PSI 产生的弱还原剂作用以合成 ATP (图7.19)。
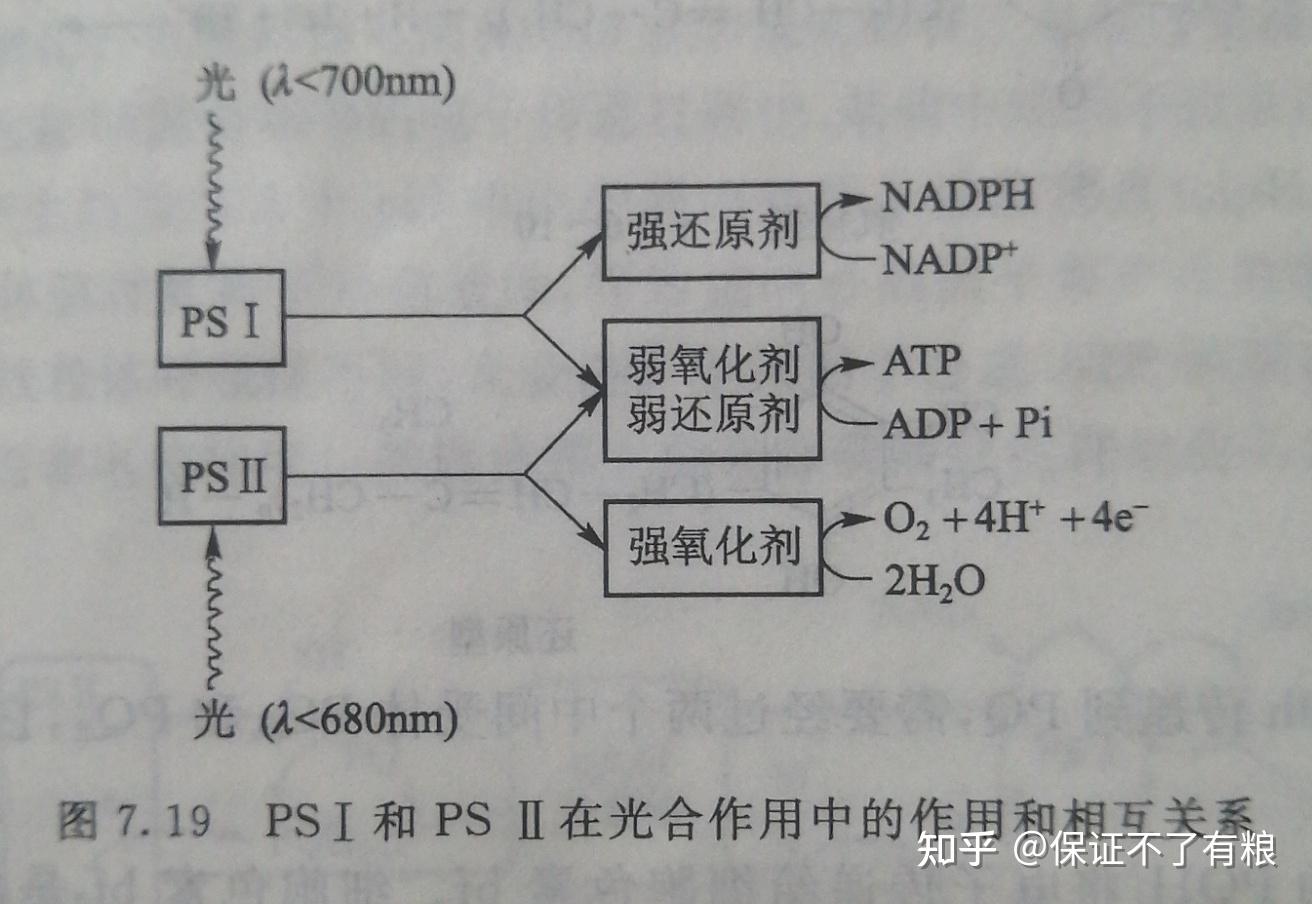
这个PSI也不是个省油的灯,里面也有“等”
(2)光反应的电子传递链(光合链, photosynthetic chain )。光反应中心的色素分子 P 吸收一个光子,即形成激发态 P*。激发态 P*的电子具有很高的能量,是良好的电子供体,因此 P*是一个强还原剂。而失去了电子的 P +,则是一个好的电子受体,是一个强氧化剂。从P*释放出来的高能电子将沿着类囊体膜中的电子传递链传递。这一个过程相当复杂,涉及两个光反应系统。光合链反应过程可分为两个阶段。
做科研的人类同胞们辛苦了!
第一阶段:在光照下, PSII 的反应中心 P680被激发,形成 P*680 。 P*680将电子传递给脱镁叶绿素( pheophytin , Ph ,为 Mg 被 H 取代的叶绿素 a ),然后再传递给质体醌,本身则变成带一个正电荷的自由基 P*680。
P*680是强氧化剂,通过 PS II 膜蛋白中的酪氨酸残基( Z )从H2O获得电子。其结果是将电子从H2O传递到质体醌。H2O和质体醌的标准氧化还原电动势分别为0.82V和0.1V,它们之间的电动势差为+0.72 V 。电子之所以能逆电动势传递,是由于P680的电子在光照激发下,具有﹣1.82V的电动势,足够克服+0.72 V 的能垒。在水裂解过程中,含Mn2+的蛋白质具有重要作用(图7.20)。
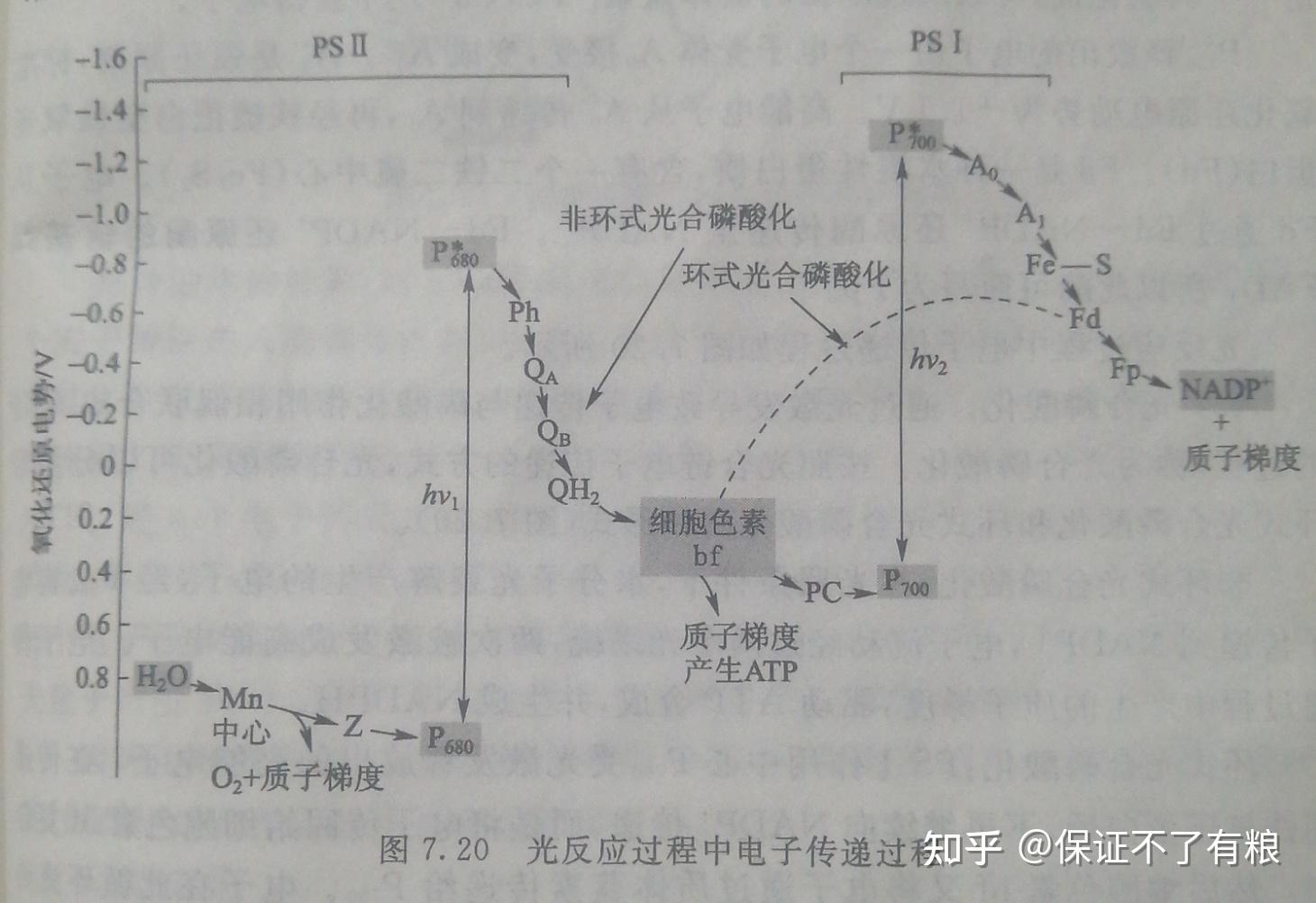
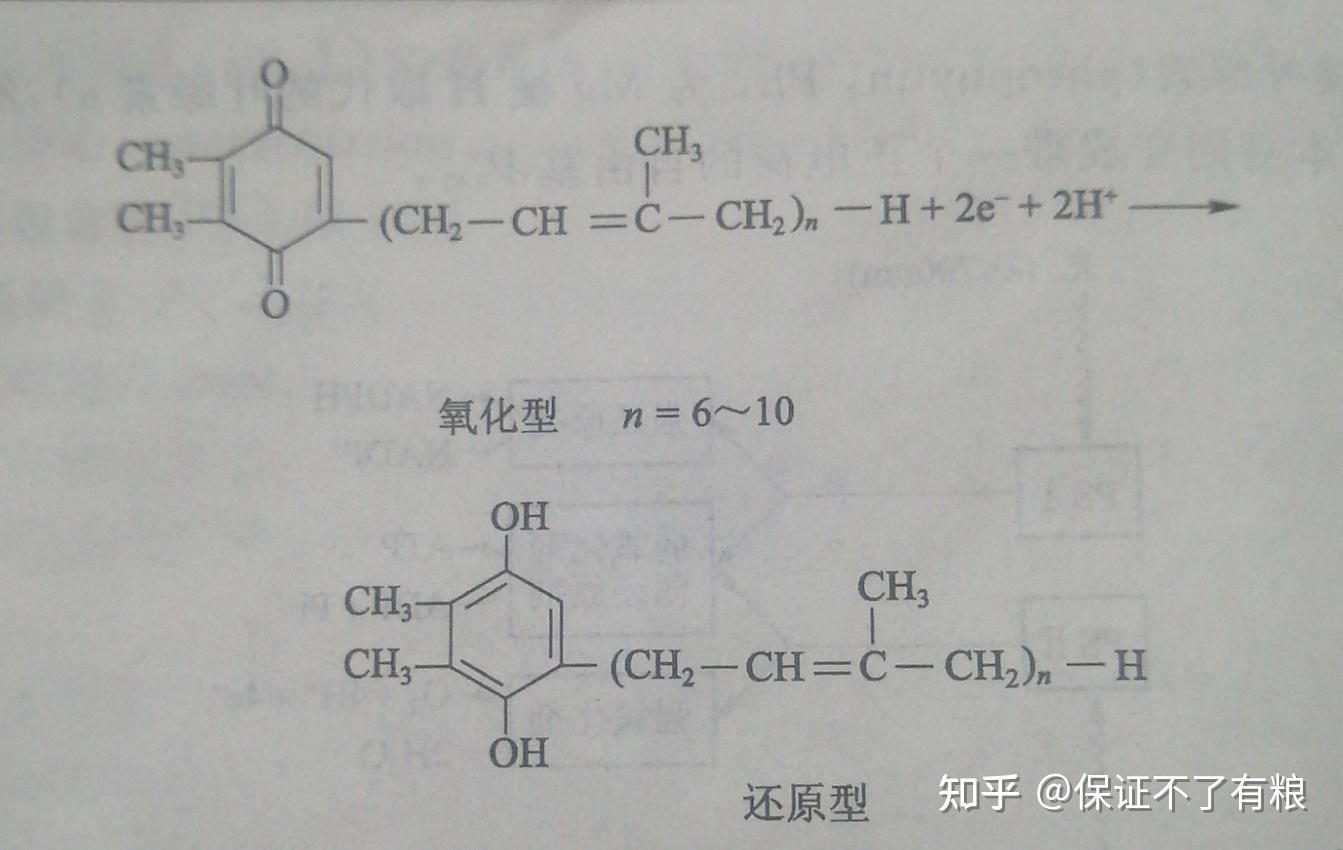
质体醌与泛醌相似,在光合链中起着电子传递中间体作用。
电子从 Ph 传递到 PQ ,需要经过两个中间受体 PQA 和 PQ B,它们是两种 PQ 结合蛋白质。
还原型的 PQH2将电子传递给细胞色素 bf 。细胞色素 bf是由细胞色素b和细胞色素f以及铁硫蛋白( Fe - S )等组成的复合物,起质子泵的作用,即在细胞色素 bf 将电子传递给质体蓝素过程中,将质子泵入类囊体膜内。

质体蓝素是一个水溶性蛋白质,它的氧化还原中心含有Cu2+。
第二阶段: PS I 经光照形成激发态P*700。P*700释放出一个电子变成P700+.它是一个弱氧化剂,可以从还原型的质体蓝素[ PC ( Cu +)]中获得电子。
P*700释放出的电子由一个电子受体 A0接受,变成 A0-, A0-是强还原剂,标准氧化还原电动势为﹣1.1V。高能电子从 A0-传递到A1,再经铁硫蛋白至铁氧还蛋白( Fd )。 Fd 是一种水溶性蛋白质,含有一个二铁二硫中心(Fe2S2)。电子从 Fd 通过 Fd - NADP+还原酶传递至 NADP +。 Fd - NADP +还原酶的辅基是 FAD ,所以此酶可简写为 Fp 。
光反应过程中电子传递过程如图7.20所示。
探索得如此透彻,生物科学家们真的辛苦了!
可是,我种个田,为什么要学这些啊,还要考试过关才行,作孽!
(3)光合磷酸化。通过光激发导致电子传递与磷酸化作用相偶联合成 ATP 的过程,称为光合磷酸化。按照光合链电子传递的方式,光合磷酸化可以分为非环式光合磷酸化和环式光合磷酸化两种形式(图7.20)。
非环式光合磷酸化:在光照条件下,水分子光裂解产生的电子,经 P680将电子传递到 NADP +,电子流动经过两个光系统,两次被激发成高能电子。电子传递过程中产生的质子梯度,驱动 ATP 合成,并生成 NADPH 。
环式光合磷酸化: PSI 作用中心P700受光激发释放出的高能电子,在传递到铁氧还蛋白后,不再继续向 NADP+传递,而是将电子传回给细胞色素 bf 复合物。然后细胞色素 bf 又将电子通过质体蓝素传递给P700。电子在此循环流动过程中,产生质子梯度,从而驱动 ATP 的合成。所以这种形式的光合磷酸化称为环式光合磷酸化。环式光合磷酸化只涉及 PSI ,并且只生成 ATP 而无 NADPH 生成。这是当植物体内需要 ATP 时选择的电子传递形式。
光合磷酸化的机理同样可用化学渗透学说来解释。在通过类囊体膜上 PSI , PS II 和细胞色素 bf 复合物等的电子传递过程中,基质中的质子被泵进类囊体膜内腔,产生约为3.5个 pH 单位的膜内外子浓度梯度。另外,由于类囊体膜对氯离子是通透的,导致膜内外因质子泵产生的电位梯度被消除。因此,与线粒体呼吸链不同,光合链产生的用于合成 ATP 的驱动力主要来自于 pH 梯度,而非电位梯度。类囊体膜上的光合磷酸化机理如图7.21所示。
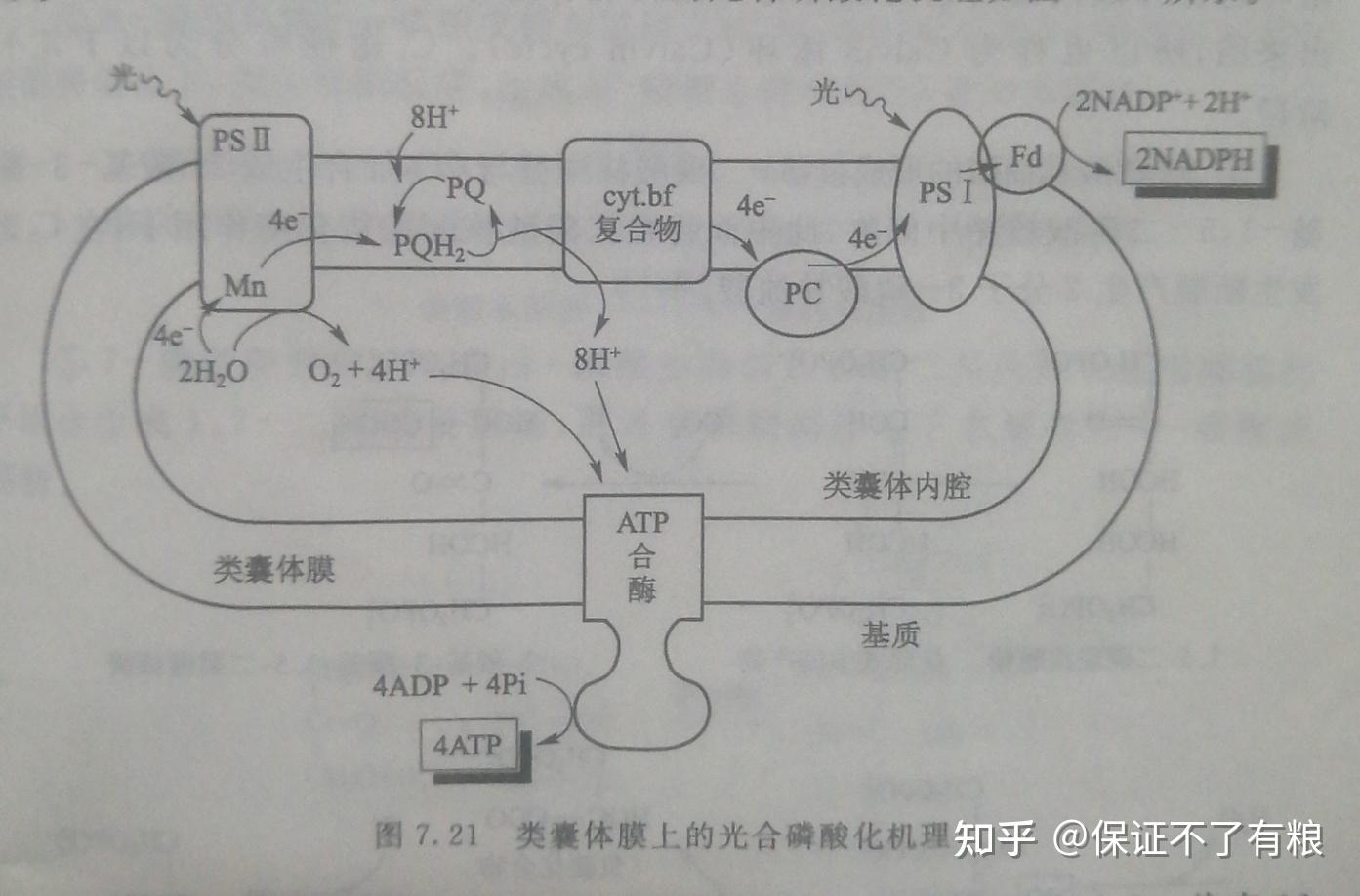
光合效率的计算:对于非环式光合磷酸化来说,每产生1分子O2,将有12个质子转运进入类囊体内腔,其中4个质子来自于水的光裂解,另外8个质子是通过4个电子的传递过程中经细胞色素 bf 复合物转运进来的。根据类囊体膜每转运3个质子可以合成1个 ATP 计算,每产生1分子O2,可以合成4分子 ATP 。经4个电子传递产生的2分子 NADPH 理论上可以合成6分子 ATP 。因此,每产生1分子O2,共可以合成10分子的 ATP 。由于从H2O到 NADPH ,每个电子至少需要2个光量子供给能量,因此,光反应整个过程的效率是每8个光量子产生10个 ATP ,即每吸收1个光量子大约能够合成1.25个 ATP 。
因为由细胞色素 bf 复合物介导的醌循环,有更多的质子转运类囊体内腔,因而环式光合磷酸化的光合效率更高。
造一个这样的机器出来应该非常非常不容易,估计比整一个windows系统出来还要难很多倍吧!
2.暗反应
暗反应( dark reaction )是指由光反应产生的 NADPH 在 ATP 供给能量情况下,将CO2还原成糖的反应过程,这是一个酶催化的反应过程,不需要光参加,所以称为暗反应。
暗反应是一个比当今地球上任何一个生化工厂的生产流程都要复杂且难以操控的过程。
这一门课,那年我反复看了三遍才过。
你们也都看累了吧,休息几天再写哈。